
How a jumping gene shaped the human skin color evolution
Two separate transposition events within ASIP, encoding agouti protein, have influenced the skin color changes during human evolution

Regular readers of my Substack would recognize my special interest in noncoding genome. Today, we explore a fascinating tale of how a mobile genetic element has been coloring the evolutionary journey of our human ancestors for hundreds of thousands of years.

One of the biggest mysteries that scientists were wrestling with during the early days was the puzzling disconnect between size of the organisms and their genomes. Larger animals have smaller genomes, whereas some plants, insects and even some unicellular organisms (e.g. Amoeba dubia) have humungous genomes. Just a couple of months ago, scientists found on the forest floors of an Australian island a plant species with the largest genome ever known to humans (Fernández et al. iScience 2024). Tmesipteris oblanceolata, a tiny fern species endemic to eastern Australia has a genome that is 50 times larger than that of ours (NYT article).
The puzzling relationship between the genome size and organismal complexity, described as 'C-value paradox', is now explained by the fact that a major part of genomes are 'graveyards' of repetitive elements containing fossilized genomes of ancient viruses. More than 45% of the human genome are repeat elements filled with transposons, aka, jumping genes. Transposons jump from one part of the genome to another and, in the process, edit, delete or create new genes, bestowing upon its host new phenotypes. They are one of the major drivers of human evolution.
Just a month ago, there was a big breakthrough in genome editing where researchers from the Arc Institute in the US and University of Tokyo in Japan uncovered the molecular mechanism through which an evolutionarily old, most simplest forms of mobile elements called insertion sequences (IS) found in bacterial and archaeal species jump across the genome(s) (Durrant et al. Nature 2024, Hiraizumi et al. Nature 2024). The breakthrough part is the realization that a component of this ancient transposition machinery was a noncoding RNA that bridges the transposon itself (bound to its one loop) with the target DNA integration site (bound to its other loop). Both the loops of this "bridge RNA" are programmable. So, by altering the RNA loop sequences, one can trick the recombinase enzyme that is part of the machinery to make large structural changes at desired target sites in the genome. You can basically cut out a large piece of DNA, or remove a piece, flip it and put it back, or even insert a whole new gene into the genome.
Okay, let's not drift into the gene editing literature. Coming back to our topic, that is, human genetics, in a recent work published in Nature Genetics, Po-Ru Loh, Steve McCarroll and team at the Broad Institute in the US report how a transposon sitting within a skin pigmentation gene ASIP has driven the evolution of skin color in humans and their ancestors.
ASIP encodes 'agouti signaling protein', a paracrine hormone produced by the skin cells. The agouti protein competes with melanocyte stimulating hormone (MSH) to bind to melanocortin 1 receptor (MC1R) in the melanin-producing skin cells. Agouti hormone blocks the activation of MC1R by MSH in the skin and slows down the production of eumelanin (black-brown pigment), skewing the balance towards pheomelanin (yellow-red pigment). Agouti protein has a fascinating literature spanning multiple species including mice and humans, worthy of a whole book. The first time I read in detail about this protein was last year when I came across a paper that reported the discovery of a structural variant in ASIP as a rare monogenic cause of morbid obesity in humans. I've told this story in a previous post. It's a fascinating one; do check it out, if you haven't already.
As was the case with many of the metabolic genes such as leptin (discovered from ob/ob mice), our knowledge of agouti gene and its function came from mouse genetics. Spontaneous mutations in agouti gene cause fur color variations in mice. This knowledge led to the successful cloning of the human agouti gene in 1994 (Kwon et al. PNAS 1994). Although candidate gene studies have reported association between genetic variations near ASIP and skin pigmentation (Kanetsky AJHG 2002), the first robust evidence came from a genome-wide association study (GWAS) by deCODE genetics in 2008 (Sulem et al. Nat Gen 2008). Studying around 5000 Icelanders, Sulem et al. identified a variant near ASIP that was strongly associated with skin sensitivity to sun, freckling and red hair, a set of phenotypes reminiscent of the genetic associations previously reported for MC1R. In the same issue, two parallel papers reported the association of ASIP locus with skin cancer risk. Although, the causal gene was pretty much clear, the none of the research teams were able to isolate the causal variant. Subsequent studies based on European population replicated the ASIP GWAS signal and also found that the risk variants increase the ASIP gene expression in the skin. However, the causal variant at this locus have remained unknown. Interestingly, later GWAS of skin color in African and East Asian ancestries failed to identify a signal near ASIP, hinting that the locus is specific to European ancestries. In line with that, a study in 2016 found the ASIP locus as one of the regions of the human genome that underwent positive selection in Europeans (Field, Boyle, Telis, et al. Science 2016).
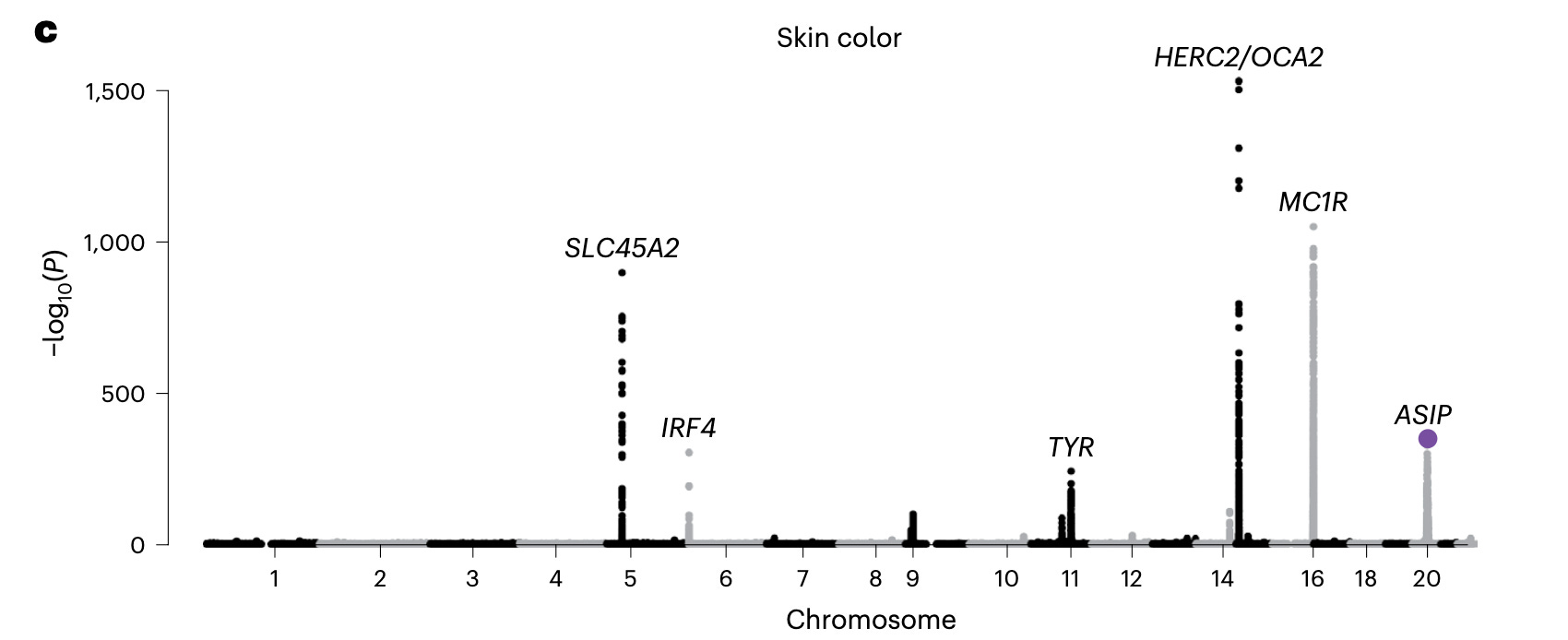
Po-Ru Loh and his team are experts in building imputation models based on common variants and using the imputation models to predict rare and complex genetic variations like rare coding variants and structural variants that otherwise can be identified only using sequencing, some only using long-read sequencing. They have been digging out fascinating variable number tandem repeats (VNTRs) in the coding and noncoding regions that were driving some of well-known GWAS associations, e.g., ACAN locus associated with height, TMCO1 locus associated with glaucoma, EIF3H locus associated with colon cancer (Mukamel, Handsaker, et al. Science 2021 & Cell 2023). Adding to this list now is the ASIP locus associated with skin color and skin cancer.
A characteristic shared by all GWAS loci previously found to be driven by structural variants was their large effect size. Given the large effect size associated with a common variant, Kamitaki et al. suspected that the ASIP locus might be hiding a structural variant underneath. So, they started digging. First they searched through the long-read sequencing data of 32 individuals made publicly availably by the Human Genome Structural Variation Consortium. One of the 32 genomes that contained the ASIP risk allele also contained a large structural variant in the same chromosome: a 3.3 kilo basepair insertion that was absent in the rest of the 31 genomes. On a closer look, the authors realized that this extra piece of DNA contained a mobile element called SINE-VNTR-Alu (SVA). SVAs are evolutionarily young and seen only in primates, including humans. They are retrotransposons, that is, they jump across the genome not through "cut and paste" approach, but through "copy and paste" approach using an RNA intermediate. So, they leave behind a copy of their own every time they jump. Further digging into the genome, the authors found that this transposon was not alone. It had a clone of its own a little further down but buried in the opposite direction, like a mirror image. Unlike the former SVA (the authors call this 'SVA F1'), which is polymorphic, meaning it is present in some humans and absent in others, the latter (SVA F) is monomorphic, meaning, it is present in all the humans. Here, things got more interesting as it appeared that not just one but two transposition events had occurred at this locus during human evolution.
To understand the functional consequence of these mobile insertions, the authors analyzed the RNA sequencing data from the GTEx database, comparing the gene expression across individuals with and without the SVA F1. Skin tissue of individuals heterozygous for SVA F1 had more ASIP expression compared to non-carriers. On closer inspection, the authors spotted one particular ASIP transcript that was decreased in expression in SVA F1 carriers as opposed to rest of the transcripts that were increased in the carriers. This aberrantly spliced transcript was almost absent in individuals homozygous for SVA F1. It turned out this aberrant splicing was caused by a splice acceptor site introduced by the non-polymorphic SVA F (SVA F1's buddy).

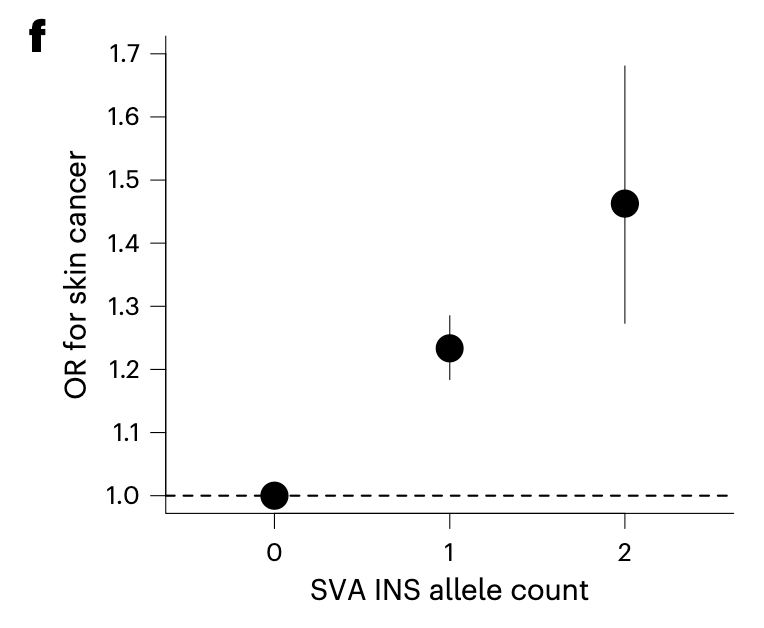
Long story short, the story unfolds like this: sometime during the evolution after the human split from the great apes but before they split from Neanderthals (~500,000 years ago), the SVA F retrotransposon jumped inside the intron of ASIP, introducing an aberrant splicing and reducing the ASIP expression. The resulting darkening of skin gave humans survival advantage, as the humans have already started losing their body hair and getting exposed to a lot of sun light. The evolutionary advantage was so strong, only the humans who carried the SVA F insertion survived, and the allele became fixed in the lineage. Then, sometime very recently during the evolution after the great migration out of Africa, a second transposition event occurred; the SVA F1 jumped straight into the same intron in the inverse direction, somehow1, releasing the ASIP expression block caused by SVA F. The resulting lightening of skin too gave some evolutionary advantage to the humans who settled over the northwestern regions of Europe. The selection pressure was strong that the SVA F1 allele rapidly rose to high frequency (7-8%) in the northwest European populations; the allele is almost absent in Asians and African populations. As always, the benefit came at a cost—increased susceptibility to skin cancer. Carrying one allele of SVA F1 increases the risk of skin cancer by 1.2 fold and carrying two allele by 1.4 fold.
Overall, it's an another remarkable work by Po-Ru Loh, Steve McCarroll and team from Broad Institute. If you think about it, the association of the structural variant itself wasn't that difficult to find. The structural variant was part of the human genome annotation. If one has looked at the genetic associations of all known structural variants with skin color or skin cancer risk, they would have found it. Yet, no one did all these years.
“The fact that the effect of such a common, large polymorphism (3.3 kb) could remain unnoticed for 15 years (even after recent advances in retrotransposon association analysis) speaks to the importance of fully integrating structural variants into genetic association analyses.”
As the authors put it, the human genomics field has ignored structural variants for a long time. There are likely a whole lot of similar evolutionary stories buried within the noncoding genome waiting to be told. Let's hope we will hear more of them in the near future.

The authors suspect that the mechanism might involve hairpin structure formation between the two SVAs in the nascent transcript, interferring the splicing process, similar to the process described for TBXT mobile element driving human tail loss evolution.
Very interesting well explained and a very informative fascinating piece with interesting insights into intron sequences, retrotransposons…..
GWAS stories definitely rocks …